C.E. Bryant, N.M.J. Rupniak, S.D. Iversen
Merck Sharp & Dohme Research Laboratories, Neuroscience Research Centre,
Terlings Park, Harlow, Essex, England
Accepted for publication June 20, 1988.
Autoaggression and stereotypies in individually housed cynomolgus monkeys were compared in a standard primate cage and an enriched playpen environment. Stereotypy and autoaggression were markedly reduced in the playpen, but reappeared on return to the home cage. Some of the various activities available in the playpen but not others engaged the animals' attention.
Key words: Foraging - Psychological welfare - Cage design
INTRODUCTION
The housing of primates in captivity is believed to promote certain types of behaviour, such as repetitive pacing or self-biting, which are normally not observed in the wild. Unlike free-ranging animals, captive primates frequently develop stereotypies (for example. repetitive rocking or cage circling [6,9,12,191) or self-directed threats or bites [1-3,9,11]. Exaggerated forms of these behaviours may be induced experimentally by total social isolation and confinement to extremely impoverished cage environments [5,6,11,15,18,231. Isolation behaviours in captive primates have been interpreted as distortions of normal social activities that become redirected towards the self and as attempts to create sensory stimulation by the animal that would normally be present in a more natural habitat [5,9,12,17]. However, it should be noted that self-directed aggression has occasionally been observed even in the wild [ 10, 14].
In macaques, these behaviours may be reduced substantially by social housing, by allowing access to foraging material, and to a lesser degree by the provision of viewing panels or mirrors [3,4,7,8,211. The apparent link between caging environment and the development of stereotypy and autoaggression in monkeys has prompted many investigators to suggest ways in which general housing conditions might be improved. Environmental enrichment devices, such as allowing monkeys access to playpens or exercise cages for short periods, puzzle boxes containing food incentives, toys such as nylon balls, and televisions or radios, have become increasingly popular in practice. However, unlike the case with socialisation and foraging, there is little empirical evidence to indicate whether isolation syndromes are in fact altered by these procedures and, if so, for how long any beneficial effects are maintained or whether primates have particular preferences for certain types of enrichment activities over others.
In the present study, we have compared the prevalence of stereotypies and self-aggressive behaviours in cynomolgus monkeys observed either in a standard laboratory home cage or in a playpen equipped with a wide range of gadgets and materials of differing shapes and textures (foraging material, viewing panel, swing, rope, ball, glove, and telephone directory). Two main issues have been addressed: 1) whether stereotypy and autoaggression are modified by transfer to an enriched environment for short periods each day and, if so, whether this effect is transient or sustained and 2) whether particular types of enrichment devices more consistently engage the animals' attention than others.
MATERLALS AND METHODS
Subjects
The subjects were six adult male cynomolgus monkeys (Macaca fascicularis) approximately 11 years of age. The animals had been housed individually in captivity for a large proportion of their adult lives and had well established cage stereotypies. Some were known to engage in mild bouts of self-aggressive behaviour. During the present study, the animals were transferred in adjacent pairs from their home cages to individual playpens located in a separate room, and their behaviour in the two environments was compared on four consecutive days each week for a 3-week period. The animals were observed at strictly the same time of day each day (between 9 am and 2 pm). The animals were fed after behavioural testing had been completed (approximately 4 pm).
Home Cage and Playpen Environments
Home Cage. For the purpose of the present study, the animals were individually housed in a single room in adjacent wall-mounted primate cages (0.72 X 1.2 X 0.6 m; Fig. 1A). Running from the front to the rear of the cage was a metal perch approximately 0.25 m above the floor. The ceiling and floor were constructed of a wire grid, the side panels of solid aluminium. sheets, and the front from vertical bars through which the animal could reach. The animals were unable to see each other, but were in full view of a separate group of monkeys housed opposite them.
Playpen. The two playpens were considerably larger than the home cage (1.22 X 1.5 X 1.05 m; Fig. 1B). As in the home cage, a metal perch ran from the front to the rear of the playpen some 0.6 m above the floor. The ceiling and floor were of wire grid construction, and the cage front was made of horizontal bars. The side panels were solid except for a perspex viewing panel located in the lower front quadrant, through which it was possible to view the adjacent playpen. The following additional items were placed in the playpens for the purpose of the experiment: a swing suspended 0.6 m from the ceiling, a nylon rope hanging from the ceiling to the floor of the playpen, a solid black nylon ball approximately 8 cm in diameter, a telephone directory, a surgical glove, and a tray placed below the grid floor of the cage containing deep woodchip litter scattered with sunflower seeds and peanuts. Damaged disposable items were always replaced before any animal entered the playpen.

Behavioural Assessment
The animals were filmed in pairs as three groups of adjacently housed monkeys using an infrared camera fitted with a wide-angle lens and a video recorder. Behaviour in the home cage was recorded for 30 min immediately prior to transfer to the playpens and for 30 min immediately thereafter. The monkeys were returned to their home cages after spending approximately 1 hr in the playpens.
Video tapes were analysed for real-time behaviours using a Philips P2000C microcomputer and software supplied by D. Harris (Royal Holloway and Bedford New College, London). Briefly, the program enabled simultaneous timing of the beginning and end of ongoing behaviours by the entry of keyboard characters previously assigned to the different activities. The following behaviours were recorded: normal locomotion, manual and oral exploration of the environment or self, resting, expression of affiliation or aggression towards other individuals, autoaggression (self-directed threats or bites), and stereotypy (repetitive pacing or other activity). Interactions with individual objects in the playpen (ball, viewing panel, rope, swing, glove, telephone directory, or foraging material) were recorded separately.
Statistical Analysis
For each 30-min recording session, the total time engaged in the above activities was calculated. Behaviours in the home cage and playpen during the 3-week period were compared by two-way analysis of variance with repeated measures. Changes in duration of specific activities during the 12-day study were determined using linear regression analysis.
RESULTS
Natural Behaviours
The duration of locomotor and exploratory activity was considerably lower (two- to fivefold) in the home cage than in the enriched playpen environment (locomotion: F = 48.28, P = 0.001; exploration: F = 283.23, P < 0.001; two-way analysis of variance; Fig. 2A,B). Exploratory activity occupied the greatest time budget ( ~ 60 %) in the playpen and was directed largely at the foraging material (see below). Linear regression analysis revealed no consistent change in either locomotor activity or exploration in the home cage during the 12-day study (r = 0.012, P = 0.92; r = 0.208, P = 0.08, respectively). However, locomotor activity in the playpen gradually decreased during this period (r = -0.294, P = 0.01), whereas manual and oral exploration was approximately doubled (r = 0.562, P = 2.78 X 10-7).
Social interactions were relatively infrequent in either environment, occupying less than 1 min. There was no difference in the duration of social behaviours in the home cage compared to the playpen (F = 1.91, P = 0.225; Fig. 2C. However, social interactions in the home cage increased towards the end of the study (r = 0.238, P = 0.04) and decreased in the playpen (r = 0.303, P = 0.01).
Inactivity or resting occupied the largest time budget of all behaviours in the home cage ( ~ 20 min). The amount of time spent at rest was consistently higher in the home cage than in the playpen (F = 507.21, P < 0.001; Fig. 2D). The duration of resting gradually increased in both environments during the study (home cage: r = 0.297, P = 0.01; playpen: r = 0.445, P = 8.9 X 10 - 5).
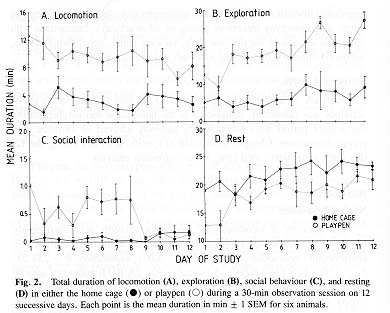
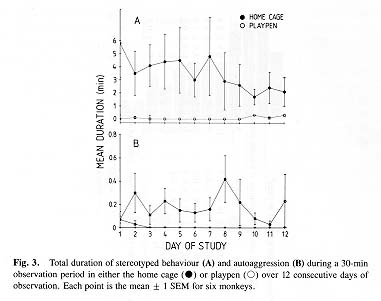
Stereotypy and Autoaggression
Stereotyped behaviours (repetitive swaying, pacing, circling, bouncing, and rocking) occupied on average 10 % of the observation period in the home cage but were virtually absent in the playpen (F = 7.02, P = 0.045; Fig. 3A). Although there appeared to be a reduction in the duration of stereotypy in the home cage as the study progressed, this trend failed to reach statistical significance (r = -0.219, P = 0.065). Similarly, there was no change in the low frequency of stereotyped behaviour in the playpen over time (r = -0.213, P = 0.073).
Noninjurious self-directed aggression (chasing and biting) was observed for an average of 11 sec during each observation session in the home cage and was rarely seen at all in the playpen (F = 7.09, P = 0.045; Fig. 3B). There was no change in autoaggression with time in the home cage (r = 0.003, P = 0.98) and a nonsignificant trend for reduced autoaggression with repeated exposure in the playpen (r = -0.230, P = 0.051).
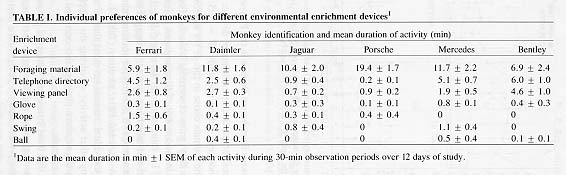
Activities Directed Towards Enrichment Devices in the Playpen
Marked preferences for particular enrichment devices over others were displayed by all six monkeys during the study (F = 195.44, P < 0.001). Notably, as a group, the animals chose to forage for approximately 10 of the 30 min during which they were observed (Table 1). There were marked individual differences in the amount of time engaged in foraging (maximum mean duration 19.4 ±1.7 min, minimum duration 5.9 ±1.8 min).
The viewing panel and telephone directory engaged the animals' attention, as a group, for an average of 2-3 min per session; other objects (swing, rope, ball, and glove) attracted little attention (Table 1). Examination of object preferences of individual animals revealed that all six animals chose to forage more than any other activity; shredding the telephone directory or viewing the animal in the adjacent playpen also occupied appreciable time budgets for four of six animals.
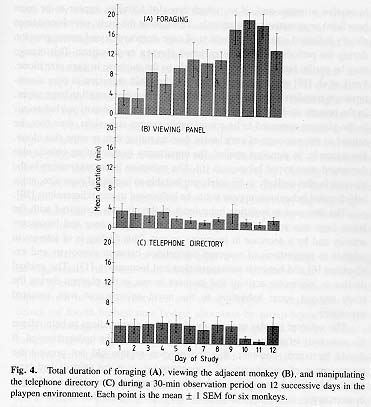
The three devices that most consistently engaged the monkeys' attention (foraging material, viewing panel, and telephone directory) were examined for any changes over the 3-week study period. On the first two exposures to the playpen, all three objects received approximately equal amounts of attention from the animals (Fig. 4A-C). However, as the study progressed, the amount of time spent by the group in foraging activity increased markedly (by approximately tenfold; r = 0.599, P = 3.33 X 10-8). In contrast, the duration of activity directed towards the viewing panel or telephone directory remained fairly constant (r = -0.220, P = 0. 066; r = -0.161, P = 0.18, respectively).
DISCUSSION
The present study examined whether exposure to an enriched environment for short periods each day is capable of modifying isolation behaviours in captive primates and, if so, which types of activities appear to be most beneficial or attractive to the animals. Throughout the study, our observations clearly indicated a marked reduction of cage stereotypies and autoaggression during the periods spent in the enriched playpen environment. This change may be partly, but not solely, attributable to the increase in cage size alone. Paulk et al. [19] observed an approximate threefold reduction in cage stereotypies on transferring individually housed monkeys from small to large cages. In the present study, the magnitude of the reduction in stereotyped behaviour in the playpen appeared to be considerably greater and might, therefore, be related to the presence of enrichment devices rather than to cage size alone. For example, in previous studies, the opportunity to manipulate objects also decreased stereotyped behaviours [6]. The reduction in autoaggression in the playpen is also unlikely to be solely attributable to increased cage size, since self-directed behaviours appear not to be influenced by cage dimensions [ 191.
The decrease in isolation behaviours in the playpen compared with the home cage was accompanied by an increase in exploratory and locomotor activity and by a decrease in inactivity or rest. This finding is of interest in relation to suggestions of response competition between stereotypy and exploration [6] and between autoaggression and locomotion [131. The gradual decline in locomotor activity and increase in rest in the playpen during the study suggest some habitation to the novel environment with repeated exposure.
The value of regular exercise in stimulating surroundings to help relieve the monotony of captive housing should obviously not be underestimated. It should be noted, however, that the playpen regime did not prevent the reappearance of stereotypy and autoaggression once the animals had been returned to their home cages. Although we observed a trend for a reduction in stereotypies in the home cage as the study progressed, this may have been due to habituation to the experimental procedures rather than to a beneficial effect of the playpen regime on behaviour in the home cage. The level of arousal of the animals might have been relatively high at the beginning of the study because of the presence of unfamiliar filming apparatus in the housing room, and this itself might have promoted stereotypic movements [171.
Habituation to the experimental situation was also suggested by the accompanying increase in inactivity in the home cage as the study continued. Moreover, the beneficial effects of the playpen on autoaggression did not persist once the animals had returned to the home cage. Similarly, Anderson and Chamove [31 found that socialisation for brief periods did not prevent the development of autoaggression in isolate-reared macaques; this behaviour was rarely observed in animals continuously housed in pairs. An important prediction from these observations would be that enrichment of the home cage itself might lead to more sustained benefits than those afforded by the more limited exposure to a playpen routine alone.
To maintain an animal's interest during the long periods spent in the home cage, particularly when socialisation is not possible, the environment should ideally be enhanced in a way that is biologically meaningful to the monkey but is not impractical because of experimental or financial constraints. Our findings suggest three such possible improvements to home cage design for future empirical evaluation. Of the many activities available in the playpen, those that consistently captured the attention of all the animals throughout the 3-week observation period were foraging, viewing the animal in the adjacent playpen, and tearing the telephone directory. Of these, the animals' overwhelming preference was to forage; however, one might speculate that some combination of these devices, such as placing sunflower seeds or other tidbits between the pages of a telephone directory, could be especially attractive to cynomolgus monkeys.
The beneficial effects of allowing captive primates to forage in reducing autoaggression has been well documented by Chamove and colleagues [7,8,21] and in the present study appeared to be equally valuable in the suppression of cage stereotypies and autoaggression. Foraging activity receives the fourth highest time budget allocation by group-housed macaques [20], indicating its biological importance to this species. The provision of foraging material appears to promote positive behavioural changes in many different primate species, both New World and Old World [7]. The increase in foraging activity observed during our study suggests that the animals learned to search for food hidden in the wood chips. The long-term behavioural effects of providing foraging material in the home cage for captive primates have not yet been documented.
The incorporation of a viewing panel between adjacent cages might provide an important means for increasing social contact between animals that cannot be housed in groups. Primates will perform operant responses in order to view pictures of conspecifics and prefer monkey stimuli over other types of pictures [22]. The provision of mirrors or viewing panels to isolate-reared macaques can reduce the tendency towards isolation syndrome behaviours [4]. However, social housing with full visual and physical access to other monkeys may not prevent the development of autoaggression and other abnormal behaviours [9].
Finally, allowing primates access to a destructible article, such as a telephone directory that can be shredded, might improve the environment in several ways, such as providing an object for tactile, olfactory, and oral exploration and possibly a substrate for bedding material. In contrast, the conspicuous lack of sustained interest shown by the animals in this study towards the other "enrichment" gadgets (swing, hanging rope, ball, and rubber glove) points to the need for careful, long-term evaluation of any putative improvements to housing conditions and allowing the animals themselves to engage in their preferred activity. Without such empirical observations, there is a risk of cluttering the environment with objects that are of little interest to the animal and have no beneficial effects on behaviour [16]. Our findings, and those of others, clearly demonstrate that different additions to the monkeys' cage are not equipotent in this respect. The possibility remains that different species of primate, or animals of an age or sex different from those examined in this study, would have different environmental requirements. Indeed, even within the macaques, it is known there are marked differences between stumptails and rhesus monkeys in the development of isolation syndrome disturbances [4,24]. Consideration of ethological factors, such as species differences in the natural habitat and time budgets allocated to different activities in the wild, may provide some valuable guidance for future research.
REFERENCES
1 . Anderson JR, Chamove AS: Self-aggression and social aggression in laboratory-reared macaques. J Abnormal Psychol 89:539-550, 1980.
2. Anderson JR, Chamove AS: Self-aggressive behaviour in monkeys. Curr Psychol Rev 1:139-158. 1981.
3. Anderson JR, Chamove AS: Early social experience and the development of self- aggression in monkeys. Biol Behav 10: 147-157, 1985.
4. Anderson JR, Chamove AS: Infant stumptailed macaques reared with mirrors or peers: Social responsiveness, attachment and adjustment. Primates 27:63-82, 1986.
5. Berkson G: Development of abnormal stereotyped behaviours. Dev Psychobiol 1: 118-132, 1968.
6. Berkson G, Mason WA, Saxon SV: Situation and stimulus effects of stereotyped behaviours of chimpanzees. J Comp Physiol Psychol 56:786-792, 1963.
7. Chamove AS, Anderson JR, Morgan-Jones SC, Jones SP: Deep woodchip litter: Hygiene, feeding and behavioural enhancement in eight primate species. Int J Stud Animal Prot 3:308-318, 1982.
8. Chamove AS, Anderson JR, Nash VJ: Social and environmental influences on selfaggression in monkeys. Primates 25:319-325, 1984.
9. Erwin J, Mitchell G, Maple T: Abnormal behaviour in non-isolate-reared rhesus monkeys. Psychol Rep 33:515-523, 1973.
10. Galat Luong A, Galat G: Consequences comportementales de perturbations sociales repetees sur une troupe de mones de Lowe Cercopithecus campbellie lowei de Cote d'Ivoire. Terre Vie 33:49-58, 1979.
11. Gluck JP, Sackett GP: Frustration and self-aggression in social isolate rhesus monkeys: J Abnormal Psychol 83:331-334, 1974.
12. Goosen C: Abnormal behaviour patterns in rhesus monkeys: Symptoms of mental disease? Biol Psychiatr 16:679-716, 1981.
13. Goosen C, Metz JAJ: Dissecting behaviour: Relations between autoaggression, grooming and walking in a macaque. Behaviour 75:92-132, 1980.
14. Grewal BV: Self-wrist biting in Arashiyama-B troop of Japanese monkeys (Macaca fuscata fuscata). Primates 22:277-280, 1981.
15. Harlow HF, Harlow MK: Psychopathology in monkeys. In Kimmel HD (ed): "Experimental Psychopathology: Recent Research and Theory. " New York: Academic Press. 1971, 203-339.
16. Line SW: Environmental enrichment for laboratory primates. JAVMA 190:854- 859, 1987.
17. Mason WA, Davenport RK, Menzel EW: Early experiences and the social development of rhesus monkeys and chimpanzees. In Newton G, Levine S (eds): "Early Experience and Behaviour. " Springfield, IL: Charles C. Thomas, 1968, 208-217.
18. Morrison HL, McKinney WT: Models of human psychopathology: experimental approaches in primates. In Frazer A, Winokur A (eds): "Biological Bases of Psychiatric Disorders." New York: Spectrum, 1977, 99-113.
19. Paulk H, Dienske H, Ribbens LG: Abnormal behaviour in relation to cage size in rhesus monkeys. J Abnormal Psychol 86:87-92, 1977.
20. Post W, Baulu J: Time budgets of Macaca mulatta. Primates 19:125-140, 1987.
21. Rosenblum LA, Smiley J: Therapeutic effects of an imposed foraging task in disturbed monkeys. J Child Psychol Psychiatr 25:485-497, 1984.
22. Sackett GP: Monkeys reared in isolation with pictures as visual input: Evidence for an innate releasing mechanism. Science 154:1470-1473, 1966.
23. Sackett GP: Abnormal behaviour in laboratory-reared rhesus monkeys. In Fox MW (ed): "Abnormal Behaviour in Animals." Philadelphia: W.B. Saunders, 1968. 293-33 1.
24. Sackett GP, Ruppenthal GC, Fahrenbruch CE, Holm RA: Social isolation rearing effects in monkeys vary with genotype. Dev Psychol 17:313-318, 1981.
This article originally appeared in the Journal of Medical Primatology 17, 257-269 (1988).
Reprinted with permission of the author.