A L Kessel and L Brent
Southwest Foundation for Biomedical Research, Department of Laboratory
Animal Medicine, PO Box 28147, San Antonio, Texas 78228 USA
Abstract
Eight baboon groups (Papio sp.) were observed for over one hundred scan samples both before and after the pro vision of structural enrichment. Additions to their home-cage included a galvanized ladder suspended horizontally by chains and a plastic drum hung from the ladder. Observations were conducted for three weeks before and three weeks after the structures were added to determine changes in space use. The baboons' age-sex class and location were recorded at 10 minute intervals over a 60 minute time period. Groups were categorized as small, medium or large for analysis. The baboons spent most of their time on the floor and the bench and this pattern did not change with the addition of the new structures. The female baboons used the new structures an average of 16.5 per cent of the observation time, the males used them 13.6 per cent of the time, and the infants used them 10.1 per cent of the time. Of the new structures, males, females and infants all used the ladder the most. Females and infants used areas that were inaccessible to males and no group size differences were found.
Keywords: animal welfare, baboon, enrichment, group size, space utilization
Introduction
In the wild, baboons (Papio sp.) spend up to 70 per cent of their time on the ground covering from 300 to over 4000ha while searching for food, but they are known to be agile climbers that spend their nights sleeping on cliffs or in trees (Altmann 1974a; Harding 1976; Hamilton 1982). Wild baboons use trees as food sources, and rocks and cliffs to avoid predation as well as using them as sleeping or resting sites (Altmann 1980; Strum 1987; Barton & Whiten 1993). Savanna baboons have been reported to travel from 5-10km (3-6miles) in one day (Harding 1976). When not traveling or feeding, baboons occupy the remainder of their time engaged in social activities or resting (Harding 1976). In captivity, the environment and environmental demands are very different than in the wild. Captive baboons do not have to search for food and water, are sheltered from predation and the elements, and may be housed in a variety of social settings such as all male groups. The baboon's natural habitat is structurally complex, while the captive environment is often structurally simple and physically restrictive.
There is a great deal of information available on baboons in the wild, but the information pertaining to enrichment for captive baboons is limited to a few abstracts and little quantitative data. Many of the techniques employed for baboon enrichment have been borrowed from studies with macaques. However, baboons respond differently than macaques to structural enrichment. Baboons are more destructive than macaques and Harris (1988) found that mate and female baboons spent more time manipulating the environment than male and female macaques (Macaca fascicularis).
Of the thousands of baboons used in biomedical research, many cannot be housed in naturalistic enclosures due to study constraints. Single housing or small-group housing is common. In studies with singly-housed female baboons Jerome and Szostak (1987) found that the baboons utilized hanging ropes, chains and bars, but foraging devices were used more consistently and social pairing was found to be the most effective form of enrichment. Choi et al (1992) also found that baboons in both single and gang housing preferred foraging devices to other forms of enrichment, although juvenile baboons spent more time on structural enrichment such as chains and a Primahedron (Double L, Garnavillo, Iowa). In an attempt to provide singly-housed baboons with stimulating natural foods, corn on the cob was fed to baboons of various ages and life histories and was found to reduce cage stereotypies (Bennett & Spector 1989). In another study, four female baboons were provided with a PVC foraging device that significantly reduced abnormal, inactive and self-directed behaviours; as well as social behaviours (Brent & Long 1995).
Providing nonhuman primates with various forms of structural enrichment has become a popular option in many zoo and laboratory settings where there may be limitations on the size of the enclosure. Physical additions to the enclosure can allow greater possibilities of vertical and horizontal space usage. Structures may provide visual barriers or hiding places as well as novel areas for play or exploration. Increasing structural diversity can also provide primates with access to better views, more secure perches, escape routes and additional exercise opportunities (Maple 1979). When changing the environment or adding structures, it is important to determine if the nonhuman primates change their space usage patterns. This information can be used to develop effective housing designs, to establish preferences and determine space usage.
Studies which addressed space utilization in other species have indicated that captive gorillas, chimpanzees and macaques prefer to be near or on vertical structures (Stynes et al 1968; Menzel 1969; Hughes & Menzel 1973; Traylor-Holzer & Fritz 1985; Ogden et al 1993; Goff et al 1994), with dominant animals claiming preferred areas (Stynes et al 1968; Hedeen 1982). The vertical structures are typically immoveable objects along edges and corners, such as walls or a fence. In a study of chimpanzees introduced to a new enclosure, Brent et al (1991) found that the chimpanzees used corners, perimeters and level, nonmoveable sites the most. Moveable objects were the last to be explored and were used most often by adult males for display purposes. In a study of space utilization in singly-caged subjects, Harris (1988) reported that baboons showed maximum usage of the front perch. Information on the use of structural enrichments for group-housed baboons is lacking. Variations in group size may have consequences concerning enrichment use. There may be more competition for preferred locations in groups with a greater number of individuals.
Although information on the behavioural effects of structural enrichment is also important, this study focused specifically on changes in space use. The purpose of this preliminary study was to determine usage of new structures by socially-housed baboon groups and whether those new structures altered the baboons' typical patterns of space usage.
Methods
The baboons used in this study were Papio hamadryas anubis, Papio h. hamadryas or hybrids. All baboons were housed in groups during the study in indoor/outdoor enclosures that measured 5.79 x 8.53 x 6.10m. Each enclosure had a cement platform, or bench, that was 1.52m off the floor, 5.49m wide and 1.83m deep. Groups ranged in size from 5 to 21 animals. The baboons were exposed to daily weather fluctuations and normal husbandry activities occurring around the facility. Enclosures were cleaned and baboons were fed their standard diet once daily. In addition, the baboons received periodic enrichment consisting of grains, fruits, vegetables or peanuts. Because the subjects were taking part in ongoing biomedical studies, the were fed different diets and individuals were moved on occasion.
Eight baboon groups of varying composition were observed for three weeks prior to the addition of structural enrichment and again for three weeks following the addition of the new structures. Each group consisted of one adult male, a number of females and possibly infants or juveniles. The baboons age-sex class and location were recorded each afternoon during scan sampling periods of 60 minutes with 10 minute intervals (Altmann 1974b). A total of 122 one-hour scan sampling periods were collected throughout the study period. The total number of scans per age-sex class and group size varied due to weather conditions and management decisions such as moving females to alternate breeding groups. In the baseline condition observations were based on the following locations: floor, bench, struts (steel support girders), the wire on the front, sides, back and roof, the curb along the side, and the roof beams. The structural enrichment consisted of a 2.31 x 0.53m galvanized ladder suspended horizontally by chains and two 250 litre (55 gallon) plastic drums with holes cut in the sides hung from the ladder. After the new structures were placed in the enclosures, the following locations were added: ladder, barrel (contacted from the floor), barrel (on top of or in the structure), and new chains.
Data were analysed with Systat for Windows Statistics Package (SYSTAT Inc, Evanston, Illinois) to determine if changes in space use were found before and after installation of structural environmental enrichment, and if space use patterns varied by group size or the subject s age-sex class. Groups were categorized as small (5 to 10 baboons), medium (11 to 16 baboons) and large (over 17 baboons), for analysis. Age-sex-classes used in analysis were adult male, adult female and infant. Juveniles were not included in the analysis due to the small and unbalanced sample size. Because group membership and the number of possible locations changed during the study, nonparametric, procedures were used to analyse the data. The Mann-Whitney U test was used in comparisons of the use of each location before and after the new enrichment structures were placed in the enclosure. The use of the new structures was tested against a null hypothesis predicting zero use, which would be expected if the structures had little or no impact on space use patterns. Comparisons between more than two groups were completed with the Kruskal-Wallis procedure. Significant findings were further analysed with nonparametric multiple comparison tests (q). Observations on infants were unbalanced for group size, so the appropriate adjustment to the multiple comparison test (Q) was made (Dunn 1964). A level of P < 0. 05 was defined as significant for all tests.
Results
The use of different areas of the baboons' enclosures did not vary in relation to group size (P > 0. 05), so this factor was not considered in subsequent analyses.
The overall analysis, to determine the changes in area use after the provision of structural enrichment, indicated significant differences in the use of the wire side (U = 64, P < 0.006). As the baboons did use the additions to their cages, the use of the new structures was significantly greater than zero (ladder: U = 8, P < 0. 003; barrel-floor: U = 12, P < 0. 007; barrel-structure: U = 20, P < 0. 039; new chains: U = 16, P < 0. 0 17).
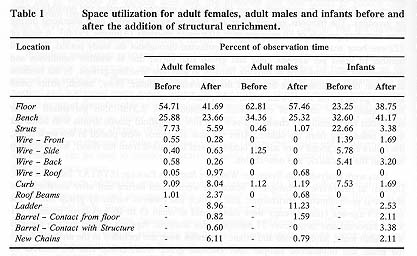
The subjects spent an average of 13.4 per cent of the data points on the new structures. The new structures were utilized by the adult females 16.5 per cent of the observation time, 13.6 per cent by the adult males and 10. 1 per cent by the infants (see Table 1).
Further analysis of the data subdivided by age-sex class demonstrated that the females used the floor, bench and wire back significantly less after the new structures were added (U = 9, P < 0. 05 for all), and utilized all of the new structures significantly more than zero (U = 0, P < 0.037). Males used the wire side significantly less after the new structures were added (U = 9, P < 0. 037), but of the additional structures the males used only the ladder significantly more than zero (U = 0, P < 0.037). Infants space usage patterns did not differ significantly before or after the provision of the new structures (P > 0. 05).
All subjects preferred the floor and bench areas, spending approximately 73.8 per cent of the data points in these locations. The ladder was the next most frequently used area for adult males and females, while the struts were the next most frequently used by the infants.
Comparing the mean levels of use of each area by adult males, adult females and infants with Kruskal-Wallis procedures, indicated significant differences in the following areas: floor, struts, wire front, wire back, curb and roof beams. Further nonparametric multiple comparisons indicated that infants spent significantly less time on the floor than either males or females (males: Q = 4.397, P < 0.001; females: Q = 2.655, P < 0.05). Infants spent significantly more time than the males on the struts (Q = 2.88 7, P < 0. 05) and wire back (Q = 4.579, P < 0.001) and significantly less time than the females on the roof beams (Q = 4.397, P < 0. 00 1). Females used the wire front significantly more than the males (q = 3.44 1, P < 0. 05) and the curb significantly more than both the males and infants (males: q = 3.747, P < 0. 025; infants: Q = 2.738, P < 0. 02).
Discussion
Wilson (1972) and others have concluded that the structures or objects present in the nonhuman primates' environment are much more important than the size of the enclosure. Perhaps if we cannot increase the amount of space allotted to our captive primate groups, we can at least impact the quality of space that they live in by providing structures that mimic features of their natural habitat (O'Neill 1988). In this study, the baboon groups were provided with large suspended ladders and hanging barrels.
The baboons spent about 13 per cent of the observation time on the new structures, with the adult males spending most of this time only on the ladder, the females using both the ladder and chains, and the infants making use of the ladder, chains and hanging barrel equally. The ladder may have been the preferred area, but access to preferred areas is often related to dominance rank (Stynes et al 1968; Hedeen 1982; Bloomstrand et al 1986; Maki et al 1989). Often times, it seemed as if one or two of the higher-ranking baboons, usually the male, would monopolize the ladders, chasing others off the ladders when approached. The ladders may have provided the best vantage points for the male baboons to view both the females in their harem, as well as neighbouring males and females. In wild baboon troops the dominant male or males have been seen as the protectors of the troop (Stynes et al 1968; Altmann 1974a).
Females and infants used a greater number of areas than the males, tending to be more spread out. Perhaps this is related to Barton's (1993) idea that subordinate animals may avoid conflict and thereby aggression, by occupying locales away from more dominant animals. There were certain areas, such as the curb and the wire front that the females used significantly more than the males. Infants used the struts and the -wire back more often than the males. All of these areas were difficult for the male baboons to access due to their larger size. Often the females would retreat to these areas to avoid male displays.
The addition of the new structures changed the space use patterns of the females the most, with decreases in the use of the floor, bench and wire areas. In general, however, all subjects still spent most of their time on the floors and bench. The preferred use of these level, non-movable areas agrees with previous reports for other primates (Brent et al 1991; Siemens-Menzies & Siemens 1992; Bettinger et al 1994). In addition, these baboons were raised in a captive setting where they had always been on cement floors, they were fed from feeders at ground level and their biscuits and enrichments were usually scattered on the floor.
While all subjects preferred the bench and floor areas, the age of the baboons may have been related to the use of the moveable objects, such as the barrels. Infants in this study were found on the swinging barrel more than the adults, who used the non-moveable structures more. In other studies, adult nonhuman primates (macaques, gibbons, siamangs, gorillas and chimpanzees) preferred stationary objects while younger animals preferred structures that moved (Maple 1979; Maple & Hoff 1982; Taff & Dolhinow 1989; O'Neill et al 1990). In a study of rhesus macaques, O'Neill (1988) found age differences in the use of perches and swings, with the older rhesus preferring the stationary perches and the younger monkeys preferring the swings and ladders. Adult chimpanzees also used level, nonmoveable structures sooner and more often, with moveable objects being the last to be explored (Brent et al 1991). One possible explanation for the younger animals preference for mobile objects may be their higher levels of activities such as play and locomotion (O'Neill et al 1990). Infants and juveniles may have used the barrels in this study as play objects, as a place to hide or as swings.
Group size is a factor that is seldom considered in enrichment studies. Although studies have shown that aggression may increase in groups of nonhuman primates due to monopolization of enrichment items (Bloomstrand et al 1986; Bloomsmith et al 1988; Maki et al 1989), differences between large and small groups have not been addressed. There is a possibility that group size could influence utilization of enrichment, either by providing individuals in smaller groups with more opportunities to use the enrichment or by causing animals within larger groups to be excluded. In this study, group size did not effect the use of structural enrichment in baboon groups ranging from 5 to 21 animals.
It is clear from this study that the addition of simple structures to baboon enclosures altered the use of the available space and provided the baboons with a greater selection of perching locations. Patterns of space use varied by the age and or sex of the subject, probably due to differences in body size, dominance status or activity level. Further studies incorporating behavioural measures and a larger sample of infants and juveniles would help to clarify age differences in space utilization, the specific uses of the various structures (as hiding places, escape routes or perches), and differences in space use due to husbandry routines or dominance status. In addition, follow-up studies would be beneficial from a management standpoint, as they could provide information on the long-term use of the added structures.
Animal welfare implications
Structural enrichment has been provided to enhance the environment of a variety of nonhuman primates by supplying them with better access to vertical and horizontal space, hiding areas, perches that allow a better view of their surroundings, escape routes and additional play areas. In this study, group-housed baboons were provided with a galvanized ladder, plastic drums and chains. The baboons used the new structures for approximately 15 per cent of the observation time; the subjects' age-sex class affected patterns of space usage, but size of the study groups did not.
Acknowledgments
We would like to thank Willie Gunnels and the employees of the Department of Physiology and Medicine for the installation of the enrichment structures and the care of the baboons. Animals are maintained in a facility approved by the American Association for Accreditation, of Laboratory Animal Care and in accordance with regulations of the United States Department of Agriculture.
References
Altmann J 1974b Observational study of behavior: sampling methods. Behaviour 49: 227-267
Altmann J 1980 Baboon Mothers and Infants. Harvard University Press: Cambridge, USA
Altmann S A 1974a Baboons, space, time, and energy. American Zoologist 14: 221-248
Barton R A 1993 Sociospatial mechanisms of feeding competition in female olive baboons, Papio anubis. Animal Behaviour 46: 791-802
Barton R A and Whiten A 1993 Feeding competition among female olive baboons, Papio anubis. Animal Behaviour 46: 777-789
Bennett B T and Spector M R 1989 The use of naturally occurring manipulanda to improve the psychological well-being of singly housed baboons. Journal of the American Veterinary Medical Association 194: 1782 (Abstract 107)
Bettinger T, Wallis J and Carter T 1994 Spatial selection in captive adult female chimpanzees. Zoo Biology 13: 167-176
Bloomsmith M A, Alford P L and Maple T L 1988 Successful feeding enrichment for captive chimpanzees. American Journal of Primatology 16: 155-164
Bloomstrand M, Riddle K, Alford P and Maple T L 1986 Objective evaluation of a behavioral enrichment device for captive chimpanzees (Pan troglodytes). Zoo Biology 5: 293-300
Brent L, Lee D R and Eichberg J W 1991 Evaluation of a chimpanzee enrichment enclosure. Journal of Medical Primatology 20: 29-34
Brent L and Long K E 1995 The behavioral response of individually caged baboons to feeding enrichment and the standard diet: a preliminary report. Contemporary Topics in Laboratory Animal Science 34(2): 65-69
Choi G C, Canfield R W, Hall E C, Haynes D R and Rice M 1992 Environmental enrichment strategies for baboons. Contemporary Topics in Laboratory Animal Science 31 (4): 6 (PS06) (Abstract)
Dunn 0 J 1964 Multiple contrasts using rank sums. Technometrics 6: 241-252
Goff C, Menkhus Howell S, Fritz J and Nankivell B 1994 Space use and proximity of captive chimpanzee (Pan troglodytes) mother/offspring pairs. Zoo Biology 13: 61-68
Hamilton W J 1982 Baboon sleeping site preferences and relationships to primate grouping patterns. American Journal of Primatology 3: 41-53
Harding R S 0 1976 Ranging patterns of a troop of baboons (Papio anubis) in Kenya. Folia Primatologica 25: 143-185
Harris D 1988 Welfare and Housing of Old World Non-human Primates (Macaca fascicularis and Papio sp.). UFAW Animal Welfare Research Report No 1. Universities Federation for Animal Welfare: Potters Bar, UK
Hedeen S E 1982 Utilization of space by captive groups of lowland gorillas (Gorilla gorilla). Ohio Journal of Science 82(l): 27-30
Hughes G H and Menzel E W Jr 1973 Use of space and reactions to novel objects in gelada baboons (Theropithecus gelada). Journal of Comparative and Physiological Psychology 83: 1-6
Jerome C P and Szostak L 1987 Environmental enrichment for adult, female baboons (Papio anubis). Laboratory Animal Science 37: 508-509(3) (Abstract)
Maki S, Alford P L, Bloomsmith M A and Franklin J 1989 Food puzzle device simulating termite fishing for captive chimpanzees (Pan troglodytes). American Journal of Primatology (Supplement 1): 71-78
Maple T L 1979 Great apes in captivity: the good, the bad, and the ugly. In: Erwin J, Maple T L and Mitchell G (eds) Captivity and Behavior, Primates in Breeding Colonies, Laboratories, and Zoos pp 239272. Van Nostrand Reinhold Company: New York, USA
Maple T L and Hoff M P 1982 Gorilla Behavior. Van Nostrand Reinhold: New York, USA
Menzel E W 1969 Chimpanzee utilization of space and responsiveness to objects: age differences and comparison with macaques. In: Carpenter CR (ed) Proceedings of the 2nd International Congress of Primatology (Atlanta, Georgia, 1968) Volume 1: Behavior pp 72-80. S Karger: Basel, Switzerland
Ogden J J, Lindburg D G and Maple T L 1993 Preference for structural environmental features in captive lowland gorillas (Gorilla gorilla gorilla). Zoo Biology 4: 381-395
O'Neill P 1988 Developing effective social and environment enrichment strategies for macaques in captive groups. Lab Animal 17(4): 23-36
O'Neill P L, Price C and Suomi S J 1990 Designing captive primate environments sensitive to age and gender related activity profiles for rhesus monkeys (Macaca mulatta). In: American Association Zoological Parks and Aquariums (AAZPA) Regional Proceedings pp 546-551
Siemens-Menzies A and Siemens B 1992 Comparison of living space utilization by chimpanzees in two zoological exhibits. In: Proceedings ChimpanZoo Conference, not paginated
Strum S C 1987 Almost Human: A Journey into the World of Baboons. Random House: New York, USA
Stynes A J, Rosenblum L A and Kaufman I C 1968 The dominant male and behavior within heterospecific monkey groups. Folia Primatologica 9: 123-134
Taff M A and Dolhinow P 1989 Langur monkeys (Presbytis entellus) in captivity. In: Segal EF (ed) Housing, Care, and Psychological Well-being of Captive and Laboratory Primates pp 291-304. Noyes Publications: Park Ridge, USA
Traylor-Holzer K and Fritz P 1985 Utilization of space by adult and juvenile groups of captive chimpanzees (Pan troglodytes). Zoo Biology 4: 115-127
Wilson C C 1972 Spatial factors and the behavior of nonhuman primates. Folia Primatologica 18: 256-275
This article originally appeared in Animal Welfare 5: 37-44 (1996).
Reprinted with permission of the publisher.